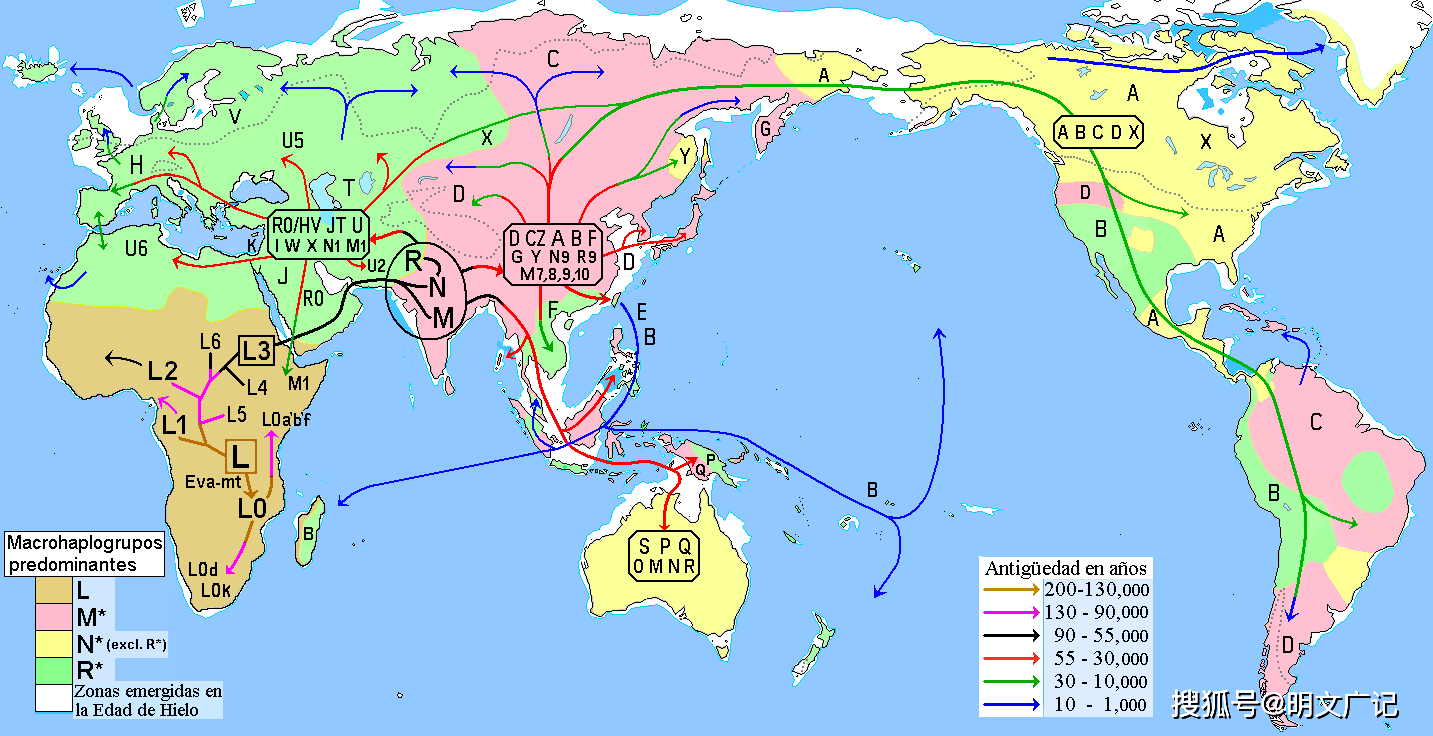
相信大家对下面这个图形并不陌生,一看就知道是说明现代人(注:现代人即现代智人,是全世界所有当代人的祖先)20万年前在非洲产生,5、6万年前从非洲走出来,迁移到世界各地并代替了当地原始人类,所以非洲现代人是现在全世界所有人类的祖先。
下面的这张迁移图还带有线粒体基因序列单倍型(mtDNA,母系遗传),内容更详细。
出非洲说的现代人类迁移路线图(带有线粒体基因类型)
按照常理,如果非洲现代人是全世界人类的祖先,并且代替了被迁移地区的古人类,说明非洲人类及优先迁移地区的人类是优先进化的,是他们把他们已经具备的文化、技术、能力带到被迁移的地区,而且代替了被迁移地区的古人类。所以迁移进来的古人类在文明、文化、能力等方面应该是优于被迁移地区的,而未迁移出来的当年的现代人群体也自然是具有同等水平的,所以原迁移地区应该优于、至少是等于被迁移地区的水平。
换句话说,在20-5万年前的现代人出现的时代,古非洲地区的人类心智能力和文化技术水平应该优于同时期的所有其他地区,先迁移的欧洲地区应该优于后迁移的亚洲地区,至少在迁移的那个时期应该是这样的。 当然,人们进入了欧亚的较严酷环境可能会锻炼了心智和其它适应能力,导致了欧亚人在这些能力方面高出同时代的非洲人,那至少欧洲因为是先迁移的地区,欧洲人理应发展出较东亚人更高的心智能力和文明水平。
大家还记得1944年美国学者莫维斯创造的“莫维斯理论”吧,西方考古学界曾认为东亚地区人种落后,文化落后。但是在中国考古事业发达以后,大量的考古和历史研究表明,上古时期以前,中国地区的文化技术水平都是明显大大优于非东亚地区的。详见明文广记文章:《一块石头也能被西方拿来歧视中国:最新考古发现再添反驳力证》、《又发现比良渚还大的史前古城遗址:中华文明是一元还是多元?》
按照出非洲说,东亚地区是比较末端的现代人被迁移地区。但是无论从历史记载还是考古研究来看,从20万年前到中国的上古时期,无论是旧石器时代,还是新石器时代,原始人的石器、骨器、陶器、玉器、青铜器、铁器、装饰品、渔猎、农耕、养殖、艺术、及其他文明文化等等,按照同期比较,都明显的表现出东亚地区优于非东亚地区的水平,无论是中亚西亚南亚,还是欧洲非洲,都是如此。
一方面按照出非洲说,欧洲文明水平应该优于同期的东亚水平,一方面实际上东亚文明水平优于同时期的欧洲水平。为什么会出现这种悖论呢?事出反常,必有妖孽。本文就来分析一下原因到底在哪里。
一、出非洲说DNA进化树及其问题
可能有人会以为出非洲说是根据分子生物学理论、根据古人类DNA基因序列产生的年代顺序排出的DNA进化树,所以是科学的理论。但请注意,出非洲说其实是一种假说,并且是一种来自分子系统发生学的假说,而这类假说在分子系统发生学业内是公认不能被当成是确定无疑的,除非有其它独立证据的验证。
出非洲说有DNA进化树不假,但那不是根据古人类DNA基因序列排出来的谱系,绝对不是!正因为不是,所以它缺乏真正的独立验证,只是一种不确定的假说!是根据当代人的DNA来推理古代的分化事件,既然是推理,就会有推理的逻辑和前提条件,而问题就是,这个逻辑和条件真的成立吗?
出非洲说的DNA进化树是有前提条件的,首先它是依赖于"分子钟假说"和"中性理论"建立的,其次是根据当代人的DNA基因序列分布情况,依据一定的假设前提建立的DNA进化树。
所以出非洲说的DNA进化树正确与否,首先它的假设条件"分子钟假说"和"中性理论"必须是正确的;其次把这个DNA进化树应用于古人类的DNA去检验,看看古人类化石的年代顺序与DNA进化树表示的DNA基因序列先后顺序是否吻合,只有都吻合了的才能证明这个DNA进化树是正确的。
而事实情况是怎么样的呢?让我们来看一下实际情况。
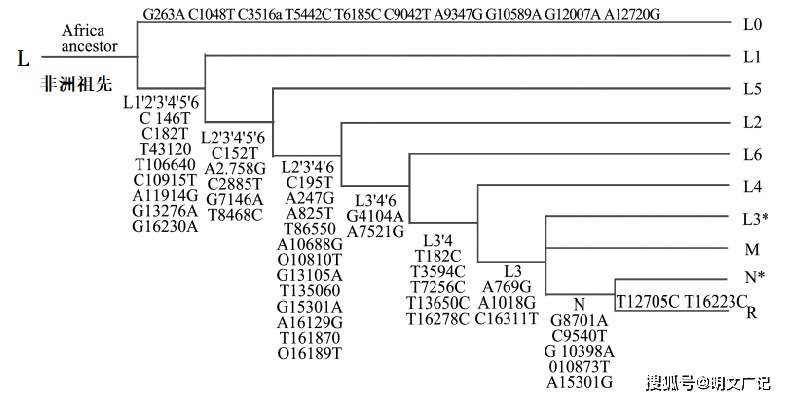
出非洲说是一种非洲单地起源说。出非洲说认为,最早在非洲进化出一批和现代人关系最近的祖先人群,并且已经表现出了全部或至少绝大部分现代人群携带的体质学特征。这个祖先人群大约于 20 万年前起源于非洲,开始时带有非洲的地域特性,随后5、6万年前从非洲向外扩散,并且最终构建出如今欧亚大陆各地区带有不同特征的人群。
在此过程中,这些出生于非洲的祖先人群完全取代了欧亚大陆上生活的直立人,发生的基因交流非常少。欧亚大陆地区的古老人种均未能成功进化成现代人,他们的后代要么自然灭绝了,要么被从非洲迁徙而来的现代人取代了,最多只对当地的现代人贡献了一小部分基因。
根据出非洲说的假设和逻辑建立的DNA进化树(进化即图谱、谱系,在本文中为同一含义)如下所示:
出非洲说的DNA进化树最开始是按照这个逻辑链条建立的:1、遗传等距离——2、分子钟假说——3、中性理论——4、遗传多样性最高的人种进化时间最长,因此这个人种的祖先就是所有人的祖先——5、用DNA序列相似性找血缘关系确定DNA序列的上下游关系。(后来还有用其它逻辑构建的出非洲进化树,比如通过外群定树跟方法,但也同样需要假定中性理论成立。)
接下来详细介绍一下出非洲说DNA进化树的建立过程。
1、遗传等距离
1963年美国生物化学家马戈利来希首次发现了令人震惊的遗传等距离现象。
遗传距离,也叫遗传多样性,是指两个物种因为基因序列不同而体现出来的基因差异。这个差异一般是指两个物种群体差异的平均值。
例如,两个物种的同源蛋白(名称一样来自同一祖先,然后分开积累突变导致序列差异),假设长度为100 个氨基酸。他们俩比对时,部分基因序列是一样的,部分基因不一样,不一样是因为突变积累。如果是20个位置不一样,则遗传距离就是20/100=20% 或 0.2。
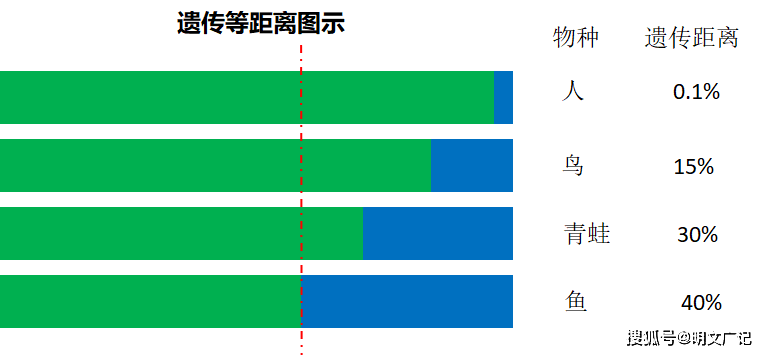
遗传等距离是指任何一个低等生物与其他多种高等级的生物相比,这些高等级生物虽然各自也有不同等级的区分,但与低等级生物的遗传距离是大致相同的,即基因序列差异的比例是相同的。这是因为这些生物间的遗传距离只能取决于被比较的这个低等生物的遗传变异,所以是相同的。这就是遗传等距离。
遗传等距离现象可以被遗传多样性上限理论完美解读。下图中绿色部分为各物种不可变异的基因序列,蓝色部门为可以变异的基因序列,复杂高级物种自身可以容忍的最大限度的变异范围要小于低等物种,因为高等物种需要更加精密的基因组作为建构零件。据此可计算各物种本身的可容忍的遗传变异范围或遗传距离的上限,一个物种本身的遗传距离上限指的是该物种类里遗传差异最大的不同种或亚种之间的距离,见右侧数据。人、鸟、青蛙都是从鱼这个物种进化来的,都带有鱼的基因序列,所以各物种红虚线左侧部门的基因序列与鱼都是相同的。
所以人、鸟、青蛙与鱼相比,基因序列不同的部分都是40%,所以他们与鱼相比,遗传距离都是40%,即人、鸟、青蛙与鱼的遗传距离都等于鱼本身的最大或上限遗传距离,鱼本身的最大遗传距离指的是鱼类里差异最大的不同鱼种之间的距离。即高等级生物与低等级生物相比,他们之间的遗传距离都取决于低等级生物本身的最大遗传距离。
注意比较对象的不同,如果人与青蛙相比,遗传距离 就是30%,所以遗传距离还是人与青蛙近,与鱼远,另外,鸟与青蛙比,遗传距离也是30%,也是鸟与青蛙近,与鱼远。
2、分子钟假说
马戈利来希发现遗传等距离现象后,用分子钟假说来解释该现象。分子钟假说认为不同物种的进化速率大致相同。注意这个假说与上述遗传多样性上限理论完全对立,一个认为遗传距离没有上限,一个认为有。
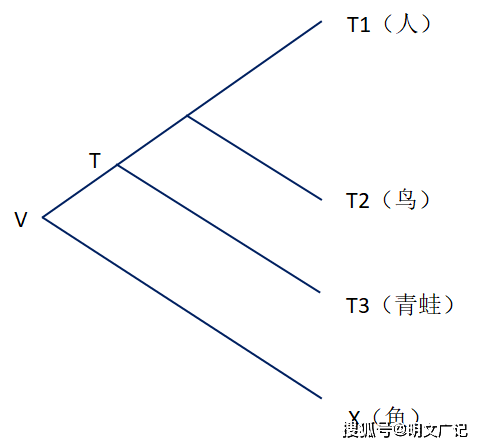
遗传等距离结果和分子钟对它的解释如下图所示。 这个示例把三种不同复杂等级的四足动物物种(T1–T3;人类、鸟类、青蛙)与另外一个低等物种(X;鱼)进行比较,这些四足物种具有最近的共同祖先 (T)。 物种 X 是外群物种并且是与物种 T1-T3 具有较远的亲缘关系,而T1-T3这些物种是群内物种。物种 T1-T3 的进化谱系在同一点 V 与 X 的谱系分离。此外,物种 T1-T3 是进化过程的产物,该过程自 V 以来已经运行了完全相同的时间。
从T这个共同祖先点开始,人、鸟、青蛙各自都进化了同样长的时间,化石证据显示是3.5亿年,它们共享一个共同的最近祖先V。同时,他们与鱼在V这个点分开,他们各自与鱼分开的时间就是相同的。根据前述遗传等距离现象,他们与鱼的遗传距离相同,进化时间都相同,进化速率等于遗传距离除以进化时间,所以他们的进化速率也相同。
如果当我们将同源的鱼蛋白质与来自不同其他三种动物(人、鸟、青蛙)的蛋白质进行比较时,如果这些动物与鱼的遗传距离是相等的(遗传等距离现象),那么这些不同动物差异积累的进化速率就应该是相似的 (T1–T3)。这就是分子钟,即不同物种的进化速率是大致相同的。分子钟假说就是说不同物种的进化速率大致相同,因此分子钟假说是来自遗传等距离的启发,是把距离假定成了与时间一直是成正比的线性距离,但这个假定已经被多重数据和遗传多样性上限理论证伪。
遗传等距离现象的发现,使人们注意到了遗传距离与进化时间的相关关系:只要是某两个物种与某一第三个物种的分离时间是相等的,则它们与第三个物种的遗传分子差异就是相等的。这就是分子钟假说,即认为不同物种的进化速率大致相同。
3、中性理论
分子钟的高速度直接给了木村资生灵感,让他提出大进化中的中性理论。他认为绝大多数物种间的DNA序列差异是由中性突变加上随机漂移造成的,进而推出分子进化速率以世代时间为单位是恒定的。
中性假说从分子层面上对进化动力提出了见解。木村资生提出, 进化的动力来源于分子层面上中性或者近中性的突变,所以外界环境这些无关痛痒的突变变化不会产生影响,而这些所谓中性变异也对生物适应环境不会产生影响。它颠覆了达尔文的进化动力——非中性突变和自然选择。
中性假说更加倾向于得出一个完全随机、绝对偶然的进化历程。既然中性理论中的这些突变仅仅是从分子层面上加以考虑, 自然就会认为:同类的分子, 不论在何种生物中, 都将是拥有一样的突变速率。而这正是分子钟假说的概念。其实中性学说与分子钟假说是循环论证,二者含义基本相同,中性学说基本就是为了解释分子钟而提出的,木村资生认为分子钟是他学说的最强证据,这是犯了明显的循环论证的错误。
4、遗传多样性最高的人种进化时间最长,因此这个人种的祖先就是所有人的祖先
按照分子钟假说,DNA序列变异与进化时间是成线性关系的。进化时间越长,DNA序列差异越大,即遗传距离越大,遗传距离(即遗传多样性)与进化时间成正比。那么遗传多样性最高的种群意味着进化时间最长或最先出现。这个分子钟逻辑就是出非洲模型最初建立时所依赖的逻辑。
后来,研究者还采用了另外一个方法逻辑来构建了出非洲模型,这就是所谓外群定树根方法。与外群物种(黑猩猩)DNA序列最近,意味着带有更多的祖先型变异基因,最接近外群的DNA序列,被认为是人类祖先所带有的DNA。这个逻辑貌似合理,但假定了一个位点只能突变一次,不能有回复突变或独立突变 (回复突变指的是从A变到B又变回A)。这也是需要中性理论成立才有可能。
非洲起源说将组外物种黑猩猩作为与人类最相近的物种,即人种与黑猩猩拥有最近的相同祖先,人类与黑猩猩是从相同的祖先那里分离开来的。
非洲南部的桑人(或称布须曼人)恰好是人类遗传多样性最高的人种,并且与组外物种黑猩猩和尼安德特人序列最近,因此被认为是现代人的祖先族群。
出非洲说的分子模型的建立依赖于分子钟假设和中性理论,其基本逻辑是把DNA序列的树根定在遗传多样性最高的种群里,或与组外物种DNA序列最近的种群里。而这里的祖先型变异基因指的是与黑猩猩相同的变异基因,恰好非洲南部的桑人符合这个要求,非洲桑人遗传多样性又最高,因此,业内把非洲桑人的DNA序列单倍群作为DNA进化树的根部型。
根据非洲桑人内部的遗传多样性最高得出现代人出非洲的结论。根据出非洲说,所有现代人都来自一个约20万年前的老祖母,她很可能是个非洲人,因此"出非洲说“也被称为夏娃理论。
非洲桑人,亦称布须曼人,非洲南部一支原住民民族。桑人主要生活在博茨瓦纳、纳米比亚及南非共和国的西北部,目前只有6万人。布须曼人的皮肤并不像常见的非洲黑人那样黝黑,而是呈黄色或黄褐色,面貌特征接近亚洲黄种人。来自葡萄牙的早期探险者初次遇到布须曼人时,曾误以为他们是中国人的后裔。他们有自己的语言,却没有文字。其语言非常独特,许多发音靠舌尖与口腔唇齿摩擦而成,这种发声方法产生于人类的发音器官还没有完全形成之时。
5、用DNA序列相似性找血缘关系确定DNA序列的上下游关系。
一般来说或按照分子钟假说,DNA序列相似性被认为是反映了血缘关系或分化时间,DNA序列越近则血缘关系越近,种群分化时间也越短。所以从树枝部的DNA序列开始,找到相近的DNA序列,一级级找下去,就确定了树根部在非洲的出非洲说的DNA进化树。注意,DNA序列相似性也被认为是反映了功能或生理相似性,但这个含义被出非洲说武断地无视了。
人类迁移、分化的过程也反映了DNA序列进化的过程,所以看起来DNA序列的排列形成了迁移路线图。
以上就是出非洲说DNA进化树建立的逻辑过程。
二、出非洲说DNA进化树的错误和原因分析
出非洲说的DNA进化树是在分子钟和中性理论假设的前提下建立的,虽然偶然会得出一些与化石和形态证据吻合的结果, 但更经常的是矛盾的结果。
1、非洲起源说DNA进化树模型无法通过古人类化石DNA验证
非洲起源说DNA进化树模型甚至无法满足最基本的自洽性的科学模型要求。古人类化石DNA才能真正见证DNA序列事件,才是最有发言权或说服力的。令人惊讶的是,业内一直未用古人类DNA来验证出非洲说,这是因为出非洲说的DNA进化树模型过不了古人类DNA检验这一关。把出非洲说的DNA进化树用于古人类化石DNA时,会发现DNA序列上下游关系与化石年代关系是颠倒的。
举个例子说明。在出非洲说的线粒体进化树模型里,R和N处于进化树的下游。但是古人类化石的DNA测序的结果却正好相反。
目前发现的最古老的现代人祖先共3个分别为出土于西伯利亚距今45000年的 Ust-Ishim,来自中国北方距今40328年的田园洞人,以及发现于意大利Riparo Bombrini距今39805 年的Fumane2。这三个人线粒体都属于单倍群R,其中田园洞人属于单倍群R下游的亚单倍群 B,Fumane2 更接近单倍群R下游的亚单倍群HV或者H,或者说是 NCBI中的现代人KP34013 和 KF523402。只有最古老的Ust-Ishim,不出所料的表现为是最原始的分化最少的单倍群R0。
通过对目前已有的相对年轻,距今在 39500-30000 年间的古代样本进行分析,发现只有两例属于单倍群N的个体,分别为位于罗马尼亚距今39500年的 Oase19和发掘于蒙古距今34425年的 Salkhit。
目前发现的最早的现代人祖先古人类化石单倍群为R,基本处于出非洲说DNA进化树模型的最下游。目前还发现一些古老的欧洲DNA,单倍型是N,年代类似Ust-Ishim。
这些最古老的现代人化石的单倍群R和N,反而处于出非洲说DNA进化树的最下游,说明这个进化树一个颠倒的排序,这个DNA排序是错误的。
2、北非和欧洲古人类化石DNA问题
按照出非洲说,非洲现代人迁移到世界各地,那么欧洲人的古DNA应该与非洲人更接近才对。
但事实是,北非的当代人基本是白人特征高于黑人。古DNA显示,15000年前的摩洛哥Taforalt古人也是欧亚DNA占多数,明显体现欧亚人进入非洲是更常见的事情。另外,45000年前的欧洲古人Ust-Ishim和Bacho Bira Cave,都没有带有非洲人的基因,反而是带有东亚人或南亚人的基因。如果按照出非洲说,早期欧洲现代人应该主要是非洲人基因才对。而事实上古北非人是带有欧亚基因,古欧洲人都是带有亚洲人基因。
这也说明是亚洲人进入了欧洲,欧洲人进入了北非。而不是现在的出非洲说DNA进化树排列的那样,非洲人进入了欧洲和亚洲。
3、非洲现代人起源年限问题
目前发现的最古老的完全现代人化石位于中国南方道县,距今 8 万至 12 万年,而在非洲尚未发现 2万年前的完全现代人化石。(道县遗址中的化石只有牙齿部分,无法确定其他部分是否含有现代人的特征。)
根据出非洲说理论,最早的现代人应该来自遗传多样性最高的撒哈拉以南地区,具体就是在非洲西南部纳米比亚和南非的边界地区,但最早的非洲现代人化石基本都是在东非和北非,这也从一个角度否定了出非洲说所认为的遗传多样性与人类遗传年限线性相关的关系。
根据出非洲说的分子钟计算结果,非洲现代人起源于20万年前。事实上,尽管在非洲和亚洲各个地区的大量遗址中都发现了带有部分现代人特征、距今310000-80000 年的古代人遗骸,但至今为止,考古学家仍没有找到一例早于4.5万年的携带所有现代人特征的人类遗骸。这是出非洲说的又一漏洞。
4、分子钟假说的进化时间计算问题
根据分子钟假说,基因变异与进化时间呈线性关系。所以遗传多样性越高的,意味着进化时间越长,也即是这个物种的祖先。
在计算物种分支时间时,其结果往往与现实数据不吻合。这是由于分子钟将极限距离草率地等同于线性距离,即与时间成正比,但这个分子钟假说隐含的假定是错误的。
在距今 34000 -35000 年的古人中发现了三个携带单倍群 M的个体,包括两个 Belgium 的Goyet Q116-1和 Goyet Q376-3,以及一个发现于法国 La Rochette 地区,距今28000年的个体54.1。
在出非洲说中,根据分子钟计算结论,N* 最早出现在5万年前的。单倍群M在出非洲说DNA进化树模型中被认为是单倍型 N的姊妹单倍型。所以单倍群 M 罕见于35000 年以前的而多见于 35000 年以后的古人类化石中的这种现象,明显不支持出非洲说的结论。M出现的时间计算是错误的。
5、中性理论不中性的问题
中性理论是把遗传变异大部分看成是中性的假设,而不是一个实证后的结论。这个理论最初是来自少数几个序列比对的数据,今天是海量基因组时代,中性理论已经被时间和数据推翻。过去60年的分子生物学功能研究已经发现大量被认为是中性的序列,其实具备生物功能性,不是中性的。古化石DNA序列分析也反驳了中性理论。
对于中性学说的解读,很多科学家提出了反对意见。首先,同一分子在不同的物种间, 其突变速率都是迥然不同的。近期研究发现,这种速率差异可以高达40倍。
观察到的大部分遗传变异是否为中性是一个有待实验证明的问题。越来越多的证据显示,只有一小部分的变异是中性的,大部分变异不是中性的,是与自然选择理论相吻合的。
6、关于人与黑猩猩的关系问题
在分子进化研究出现之前,形态分类学家和古人类学家通常是把类人猿类归为一组(Pongid),包括猩猩(orangutan)、大猩猩(gorilla)和黑猩猩(chimpanzee),而人是另外一组,人与类人猿分支时间大约是在1800万年前。
但在 20 世纪 60 年代,以分子钟假说和中性理论为基础的分子分类方法,误把上限饱和距离当成线性距离来处理,得出了黑猩猩是人类最近物种的结论,认为人与黑猩猩分支时间仅是 450 万年前(Wilson and Sarich,1969)。
这个时间被后来所发现的 700 多万年前的人类化石所证伪(Graecopithecus freybergi和 Sahelanthropus tchadensis)。因此业内现在普遍接受的人与黑猩猩分支的时间主要是由化石发现来决定的,这也就凸显了中性理论和分子钟方法的片面性。
来自德国、保加利亚和法国的国际研究团队在保加利亚奇尔潘(Chirpan) 附近地区发现了欧洲700万年前仍有类人猿存在的证据,这是属于古类人猿的一块前日齿化石。这比过去化石证据所反映的古类人猿延续时间延长了220万年。
以分子钟假说和中性理论为基础的分子分类方法,确定黑猩猩与人类有最近的血缘关系。后来的全基因组测序发现,其实只有 70%的基因组是黑猩猩与人类最近,而有30%的基因组是大猩猩与人最近,另外有1%的基因组是猩猩与人最近。重新分析人猿关系,发现黑猩猩和大猩猩与猩猩的基因序列相似性高于人与猩猩的相似性,显示人与类人猿类分开后,类人猿才分化出了黑猩猩、大猩猩和猩猩。计算显示人与类人猿类分支的时间是1760万年前,与化石发现基本一致。
7、出非洲说DNA进化树模型的祖先单倍型人数极少问题
出非洲说里DNA进化树模型的第一个现代个体或群体携带的是 A00 单倍型,一个目前只有数百人的极其小众的非洲单倍型,而第一次分化出现的单倍型是 AO-T,一个目前有 70 多亿人之众的单倍型。
一个群体中通常只是个别人发生了突变成为新单倍型的祖先,因此一个新出现的单倍型的人数多于祖先群体人数千万倍是极不正常的,只有新突变受到正选择向高级进化才有可能。但作为非洲说理论基础的中性理论是否认具有自然选择的作用的,认为进化是随机的,无方向的。
这种现象与中性理论相冲突,要么中性理论错了,要么进化树搞错了。当然中性理论错了,出非洲说里DNA进化树模型的前提条件不成立,这个进化树也要重新排列了。
而且出非洲说里DNA进化树里面的早期分支里有7个分支新突变形成的单倍型群体的人数都大大多于祖先群体的人数。若每一次中性遗传漂变的结果都是让带有新突变的个体的人数大大超过祖先群体,这显然是不合理、不现实的。
8、出非洲说DNA进化树模型中没有考虑遗传多样性上限问题
出非洲说DNA进化树模型、分子钟假说、中性理论都认为基因序列是随时间线性进化的,这其实也就隐含了遗传距离是没有上限的,基因序列是可以一直无穷尽的变异下去的。
这样不同物种的同源基因的相似性会随时间下降到没有任何相似性,这也就意味着一个基因的功能可以被随意编码,而这是完全不现实不合理的。细菌与人分离了至少10亿年,它们还有很多共享的同源蛋白,这些蛋白的相似性也较高,那么这种人与细菌的有限的基因序列差异是否达到了上限?
先验逻辑常识推理认为遗传距离或遗传多样性必须有上限,不可能无限变化。这种上限就决定了进化变异与时间不是线性关系,分子钟假设和中性理论是错误的,所以分子钟和中性理论计算出来的物种进化分支时间与实际情况经常对不上号。
根据遗传多样性上限理论,物种越优异,则其基因越精密,允许基因变异的比例越低,则遗传多样性越低;反之则反之。非洲桑人遗传多样性高是因为进化发育滞后,在基因相对不精密的情况下,允许基因变异的比例大,所以表现为遗传多样性高。而不是因为其存在祖先型基因,进化时间长造成的,分子钟和中性理论对此的解释是错误的。
一个简单的思想实验可以通俗解释遗传等距离。 如果我们能通过使用相同的某一基因序列,创造一个细菌,一条鱼,和一个人,然后让三种生物分化很长时间或大约 5 亿年,并假定他们那时还是各自与原先物种表型差异不大的物种,细菌中的一个基因会改变很多,比如 50%,而它在鱼中的对应基因也会变的较多但会低于细菌,比如说,30%,它在人类中的对应基因则会发生变化很少,比如说1%。 任何超过 50% 的变化对细菌是致命的; 任何超过 30% 的变化都会对鱼类是致命的; 任何超过 1% 的变化都是对人类是致命的。
细菌中的一个基因之所以能够变化比鱼或人多得多,而鱼变的能比人多很多,是因为人类的基因遇到了相对更多功能和生理限制。因此,细菌与人类或鱼类之间的遗传距离主要由细菌中的突变决定。 在这种情况下,细菌中 50% 的变化将解释细菌和人之间 50%的距离数值,也将解释 细菌和鱼之间的50%的距离数值,以及细菌与细菌之间 50% 的距离数值。也就是说一个简单物种细菌与高等物种人和鱼的距离,等于低等物种的最大种内遗传距离,这就是遗传等距离现象。同理,鱼的 30% 变化会决定了鱼与人距离值是30%。
相比之下,流行进化论中性理论会预测,人类和鱼都可以像细菌一样变化高达 50%,因此,他们之间的遗传距离都是50%或更多,不管是人与细菌、人与鱼,还是细菌与鱼 。
这里一个关键概念是,高等生物能容忍的变异,低等也能容忍,但反之不然。所以一个能容忍60%变异的低等物种,与一个能容忍50% 变异的高等物种的遗传距离只能是60%。如果不是这样而是不同物种的突变很少有重叠,则他们的距离就会是 60%+50%=110%。显然这是荒谬的,因为遗传距离不可能超过100%差异。
另外一种通俗解读遗传等距离是把人看成是一种鱼来看人与鱼的遗传距离。
先问几个简单问题,人是一种黑猩猩,对吗?对呀,有本著名科普书【第三种黑猩猩】,作者是普利策奖得主戴蒙德,认为人是第三种黑猩猩,第一种和第二种都生活在非洲的树林里,他们与人共享了一个最近的共同祖先,都是同一个祖先的后代。
人是一种猴子吗?当然,道理与人是一种黑猩猩是一样的,因为与猴子共享一个最近共同祖先。
人是一种哺乳动物吗?当然。
人是一种鱼吗?当然,所有陆上脊椎动物都共享了一个最近共同祖先脊椎动物鱼。
青蛙是一种鱼吗?当然是,道理同上。人与青蛙共享了一个共同祖先两栖动物,因此人也是一种青蛙。
在远古时代,人支系与鱼分开时,是在大部分鱼类还没有分化出现时,因此人与鱼的分开时间或遗传差异属于是鱼类的最大限度的,人应该是保留了所有鱼类的共性表型和基因型,但同时又积累了属于人的突变,因此人的序列与鱼的序列有高度相似系。
鱼类包括很多种鱼,其中分开时间类似于人鱼分开时间的两种鱼,就应该是代表了鱼类的最大遗传距离。因为进化时间已经很长,这个最大距离就是上限距离,超出这个距离上限就会变成不是鱼类而是其它物种了。
这个最大距离就应该是等于人与鱼的距离,因为人无非就是一种鱼,就是属于鱼类的变异最多的一种鱼,它与鱼的遗传距离不可能超过鱼类能容忍的最大限度的距离,因此只能是等于这个最大距离。超过意味着不再是鱼,但人是一种鱼。
同理,青蛙也是一种鱼,其与鱼的遗传距离也自然是等于鱼能容忍的最大限度的距离。这就会出现遗传等距离现象,既 鱼与人的距离, 等于鱼与蛙的距离,等于两种分开时间最久的两种鱼的距离,也即等于鱼类能容忍的最大遗传距离。
所有鱼的后代都保留了鱼的共性以及维持这些共性的基因序列,同时也有自己的特异序列,这些特异序列都是属于与鱼的共性不矛盾的,他们有的导致了一个后代与大部分鱼类有很大的表型差异,比如人或青蛙,但有的则没有改变后代作为鱼的表型,没有改变的原因是因为没有发生建构图纸或表观遗传编程的改变。
从以上分析可以看出,出非洲说的DNA进化树漏洞百出,不但跟古人类化石年代顺序对不上号,很多生物现象解释错误,而且其前提假定分子钟和中性理论也是存在错漏的,所以以此构建的出非洲说DNA进化树模型是错误的,出非洲说无法建立正确的、符合古人类化石年代顺序的DNA进化树。
近些年之所以出非洲说得到更多的认可和宣传,是因为出非洲说比多地区说和出东亚说更符合西方的政治正确,所以被西方接纳和广为传播。即使现在有很多西方生物学家发现了出非洲说、分子钟假说和中性理论的问题和不足,发表了现代人起源于东亚的文章,但是未能被主流舆论媒体报道传播。主流杂志如自然出版社所属杂志长期以来在人类学领域有政治优先科学的嫌疑,近期更是直接宣称不欢迎政治不正确的科学,很多正义学者公开谴责了自然出版社这种反科学的立场。
三、出东亚说DNA进化树的建立及其科学性
在各类物种的大规模测序结果逐渐得到完成和公开后,分子遗传学者黄石很快意识到分子钟和中性理论对遗传等距离的解释是荒谬的。经过深入研究,黄石发表了遗传多样性上限理论,即最大遗传变异理论,对遗传等距离及其他主要进化现象重新解读,并随后发现了遗传距离存在极限距离,即基因突变到一定极限不再导致遗传距离的增加。
遗传多样性上限理论也就是MGD理论,认为遗传多样性或遗传距离存在上限,并且大部分是处在上限饱和平衡态,不再随时间继续递增。遗传多样性理论吸取了分子钟和中性理论中的正确成分,即在小进化内可以采用分子钟和中性理论,但是遗传多样性达到上限饱和态以后,就不能用分子钟和中性理论来解释了,即此时基因进化与时间不再是线性关系了。
黄石根据遗传多样性上限理论创建了出东亚说的DNA进化树。出东亚说的DNA进化树是按照这个逻辑链条建立的:1、遗传等距离——2、遗传多样性上限——3、具备中心型特征的DNA序列为祖先型DNA序列——4、用DNA序列相似性找血缘关系确定进化树。
1、遗传等距离
上文已有解释,不再赘述。
2、遗传多样性上限
上文已有解释,不再赘述。
3、具备中心型特征的DNA序列为祖先型DNA序列
在 1983 年已经有学者利用线粒体构建了以亚洲人群为起源地的DNA进化树模型。这些学者们研究了包括高加索人、亚洲人和非洲人在内的 200 个样本,构建线粒体进化树,并得出两种迥然不同的现代人起源结果,并提出了一种判断最古老人群的方法:事先并不假定各种族人群的进化速率是否相同,而是首先假定 mtDNA 的最古老型或中心型的确定,应是基于高频率、存在于多个不同种族或存在于其他灵长类动物中。
结果发现,全部人群的 mtDNA 单倍型一共有30余种,其中只有1型不仅在欧洲人中,而且在亚洲人和非洲人中也有分布,因此认为普遍存在的1型为祖先单倍型。而1型在亚洲人中频率最高(亚洲人 69.6%、欧洲人58%、非洲人 14.9%),这说明亚洲人是最古老的,进而可以得出现代人出亚洲的结论,而这些亚洲人样本都是来自东亚,说明现代人出自东亚。
黄石构建的出东亚说的DNA进化树模型将线粒体和Y染色体进化树根部都定位在了亚洲人群中,重新独立地证实了1983年提出的线粒体根源在亚洲的论述。新模型的树根确定来自常理的推导,认为原始的单倍型应该是现代人群中存在最多的,因为基因突变属于小概率事件,非常罕见,只会在祖先人群当中的个别人中发生,因此原始祖先型的判断可以从某单倍型的分布广的特点来得出,带有其频率最高的族群就应该是祖先族群,这个推理的合理性也可以从它被两组不同研究课题组独立做出得到验证。
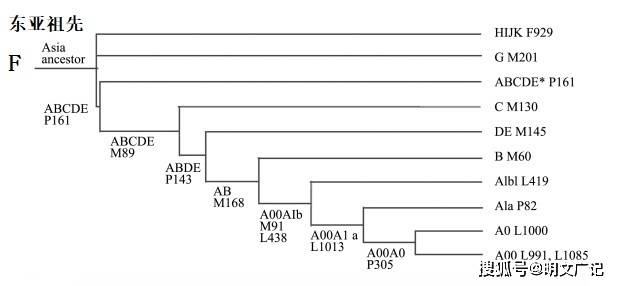
出东亚说模型认为线粒体单倍群R0是所有线粒体单倍群的祖先型,这和非洲起源说提出的 R 单倍群位于N单倍群下游的说法相左。通过研究千人基因组计划的数据,发现单倍群 R0 在现今中国南方最为普遍,这表示现代人线粒体起源地可能在中国南方。现代人出东亚说的线粒体模型也得到了古DNA分析的支持。用同样原理推导的新Y染色体进化树也把树根定在了东亚地区,认为单倍型F是祖先,而F在中国南方的拉枯族最常见。出东亚说Y染色体进化树也同样得到了古 DNA 分析的支持。英国学者通过独立研究也同样把F单倍型的起源地定在了东亚,因为欧洲人和亚洲人的常见单倍型都在F下游,这就意味着欧洲人来自东亚。
4、用DNA序列相似性找血缘关系确定进化树
从祖先型单倍群开始,按照DNA序列相近的为上下游亲缘关系,并依据遗传多样性上限理论原理,认为回复突变可以经常发生(A变成B,B后来又变成了A),就可以建立出东亚说的DNA进化树。DNA序列主要分布与出非洲说正好相反。详见下图。
注:在父系Y染色体DNA研究中,现代的汉语和藏缅语人群中都高频出现的单倍群Oa-F5,Oa-F5是上图中F下的K系的下游支系。
根据出东亚说的DNA进化树,可以描绘出东亚人群的迁移路线图,详见下图。
出东亚说DNA进化树基于科学的遗传多样性理论,不仅与古人类化石年代顺序保持一致,还可以完美地解决出非洲说存在的矛盾问题,这才是科学的人类进化路线图。出东亚说DNA进化树解决了出非洲说的诸多逻辑不自洽问题。
1、根据遗传多样性上限理论创建的出东亚模型认为线粒体单倍群R0是所有线粒体单倍群的祖先型,这和非洲起源说提出的R单倍群位于N单倍群下游的说法正好相反。
目前发现的最早的现代人化石距今45000年,线粒体单倍群为R和N,与出东亚说的DNA进化树模型正好匹配。说明了出东亚说DNA进化树的准确性。
2、所有欧亚人的Y染色体祖先是F,来自东亚,这是无争议的来自多家实验室的结论。古人类DNA测序结果表明,现代人起源于东亚,具体是出自东亚南方地区,包括今天的湖南、湖北、四川、贵州、广西、云南等地区。
3、出东亚说的分子钟计算显示人与类人猿类分支的时间是1760万年前,与化石发现基本一致。
4、古DNA分析表明,45000年前的欧洲人种Bacho Kiro 的DNA序列与东亚近,与非洲远,这说明欧洲人种出自东亚。
5、出东亚说的父系染色体根部单倍型F型分布在东亚,其下游人群普遍分布在欧亚,拥有最多的人群。祖先群体的人数应该多于支系群体的人数,基因突变的只是少数人,越下游的人数越少,这才是根部单倍型应有的现象。这证明了出东亚说的科学性。
6、黄石教授团队利用遗传多样性上限理论重新构建了灵长类动物的亲缘关系树, 重新肯定了所有猩猩是一组,而人是另外一组的常识性判断, 而这正是古人类学者长期以来的共识。
如对于人和大型猿类(如黑猩猩等)来说, 印尼红猩猩相对复杂程度低且遗传多样性上限高。因此,MGD选择红猩猩为参照来计算其他猩猩、人与该红猩猩的遗传距离。结果发现,非洲的黑猩猩和大猩猩比人跟红猩猩遗传距离近,因此, 应该把猩猩们归为一类,而人是另外一类,远在1760万年前就已经与猿类分离。
黑猩猩与人距离更近或最近,反映的完全是极限距离,就如同鸟与人的遗传距离比蛇与人的遗传距离更近, 但鸟与蛇却属于同类。这种基于MGD理论基础上的分子分类方法所得出的结论与独立演算的化石证据契合度非常高,对人猿关系重新给出了一个符合常识和化石证据的科学结论,优于基于中性理论的结论,即人与黑猩猩为一类,并仅在5~7百万年前人猿才分开。并有力支持了中国考古学者对800万年前生活在云南的禄丰古猿所作的判断,即其为人科。
出东亚说与较早提出的化石版本的东亚人本土连续进化模型是一致的,东亚人是本土连续进化了200万年,东亚地区的现代人主要源自本土的连续进化,北京猿人是中国人的直系祖先,中国有上百万年的人类史。
7、四十年间有7个独立方法和角度都得出了现代人起源于东亚的同样结论,包括:
①吴新智团队的化石证据链
②Cavalli-Sforza团队的线粒体结论
③黄石团队在遗传多样性上限理论指导下对现今DNA的分子研究结论
④黄石团队利用古DNA单亲染色体数据对出东亚说的验证
⑤Pääbo团队的早期欧洲保加利亚古人主要是东亚基因的结论
⑥Tyler-Smith团队的欧州和西亚的Y染色体来自东南亚的结论
⑦Munch团队的欧洲和西亚的X染色体可能来自东亚的结论
如此之多的独立发现都指向同一个结论,说明出东亚说是科学的结论。
中南大学遗传学黄石教授的“遗传多样性上限理论”和“出东亚说DNA进化树”被分别写进国外国内两本教科书:一本是美国教授David Bickel写的英文书《系统发育树与分子进化》;另一本是国内朱作斌教授等学者们合写的中文书《系统生物学》。书中介绍了黄石教授团队以遗传多样性上限理论为基础推导的现代人起源自东亚的理论,以及用古DNA验证东亚起源的证据。这也是世界上首次用古DNA区分不同起源假说。
美国教授David Bickel写的英文书《系统发育树与分子进化》
遗传多样性上限理论和出东亚说DNA进化树说明现代人是从东亚走出,并迁移到世界各地,通过杂交混血等方式同化了当地的古人类。所以我们也就明白了为什么远古时代东亚地区的人类心智能力和文化技术水平要高于非东亚地区,原因就在于此。
遗传多样性上限理论中的上限或最佳平衡概念, 与中国传统哲学易经中的宇宙基本规律:阴阳平衡、中庸之道的思想是一致的,它能更科学合理的解释困扰人类数十年的遗传等距离现象和生物进化问题,为分子生物学的发展作出了重要贡献,为人类遗传进化历史点亮了明灯,让我们真正的看清古人类进化发展之路。
参考文献:
东南大学出版社,《系统生物学》,朱作斌、张潇、王亮主编
2019年《人类学学报》,《古DNA的新发现支持现代人东亚起源说》,张野、黄石著
2021年《今天非洲之外的人类Y染色体全部起源于东南亚》,英国桑格研究所的克里斯·泰勒·史密斯教授等著
2023年1月31日《科技日报》,《新研究为“出东亚说”再添力证》,骆香茹著
以上一并表示感谢!
本文版权属于“明文广记”所有,欢迎分享、转载或引用,如果转载或引用请注明来源于“明文广记”!
欢迎关注“明文广记”,一起发现新奇的历史文化!








 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 照妖镜
照妖镜